В 1798 году в Лондоне впервые вышел в свет двухтомник небезызвестного английского священника и экономиста Томаса Роберта Мальтуса "Опыт о законе народонаселения". Этот же год считается годом рождения т.н. мальтузианства, которое уже два столетия висит черной тенью над всем человечеством.
Томас Мальтус, исследуя динамику изменения численности населения Северной Америки во второй половине 18-го века, заметил, что она удваивается через каждые 25 лет. Позднее, обобщив эти свои исследования и распространив их выводы на весь земной шар, он писал: "Если размножение населения не встречает никакого препятствия, то оно удваивается каждые двадцать пять лет и возрастает по геометрической прогрессии", а "средства существования в самых благоприятных условиях для труда ни в коем случае не смогут возрастать быстрее, чем в арифметической прогрессии" ([1], т.1, с.9, 162). В результате по Мальтусу создаются "лишние" люди, о которых он предрекал: "На великом пиршестве природы для него нет прибора. Природа приказывает ему удалиться, и если он не может прибегнуть к состраданию кого-либо из окружающих, она сама принимает меры к тому, чтобы ее приказание было приведено в исполнение"([1], т.1, с.12).
Как видим, приговор Мальтуса весьма жестокий. Его же последователи - неомальтузианцы, развивая теорию своего патрона, пришли к еще более мрачному выводу: наступит время, когда людям не будет хватать не только пищи, но даже "места под солнцем".
Несмотря на кажущуюся фундаментальность исследований Томаса Мальтуса, он допускал две грубейшие ошибки, не простительные даже школьнику (а ведь он был еще и членом Лондонского королевского общества - эквивалента национальной академии наук). Во-первых, он не учел, что в его время население Северной Америки увеличивалось в основном за счет переселенцев из Европы. Во-вторых, он не учел или не знал, что к 1750 году население земного шара удвоилось за предыдущие почти 750 лет.
В последующем ученые, конечно, заметили эти ошибки Мальтуса, однако, игнорировали их, потому что прогрессирующее размножение людей даже за счет только родившихся слишком очевидно. Причем, люди размножаются даже не по простой геометрической прогрессии, а по экспоненте. Так, если по расчетам демографов население земного шара к 2000 году удвоится за предыдущие 39 лет, то 9 предшествоваших этому удвоений происходили соответственно за 65, 120, 300, 1600, 2500, 4000, 25000, 60000 и 300000 лет [2]. Следующее же удвоение, если ничего кардинального не произойдет, будет всего за 15 лет, т.е. даже быстрее, чем это определил Мальтус (в скобках замечу, что, например, численность населения Ирана удвоилась за 1981-95 гг., включавшие кровопролитную Ирано-Иракскую войну, унесшую около 500 тысяч жизней молодых мужчин). Вроде подтверждается и вывод мальтузианства о том, что со временем людям не будет хватать продуктов питания - уже сейчас только от голода в мире ежегодно умирает 15 миллионов человек, что равноценно взрывам около 200 таких атомных бомб, которая в 1945г. была задействована над Хиросимой.
Как видим, мальтузианство правдоподобно. Правдоподобна также т.н. "биологическая основа" мальтузианства, постулирующая, что в благоприятных условиях и п и отсутствии естественных врагов всякий биологический вид способен на бесконечное прогрессирующее размножение. (Впрочем, в более современной формулировке этот постулат звучит и так: прогрессирующее воспроизведение себе подобных является внутренним неотъемлемым свойством живой материи). Применительно же к человеку этот постулат означает, что, поскольку люди создали себе максимально благоприятные условия и не имеют естественных врагов, то им не остается ничего другого, как размножаться прогрессирующе и неограниченно.
Правдоподобность мальтузианского постулата ввела в заблуждение многих, даже таких именитых ученых, как Чарльз Дарвин и Альфред Уоллес, которые положили его в основу своей эволюционной теории - дарвинизма. Однако, постулат этот глубоко ошибочен и не имеет никакой биологической основы. Доказательств этому существует много, из которых укажу лишь на следующие общеизвестные факты:
Первое уравнение модели Лотки-Вольтерры, с которым мы будем иметь дело, выводится следующим образом (см., например, [4]).
Если x = x(t) есть численность особей популяции жертв в системе "хищник-жертва" в момент времени t, а N = const - скорость размножения единичной особи той же популяции, то при отсутствии хищников скорость изменения числености особей всей популяции будет:
![]()
Проинтегрировав это уравнение, получим:
x = С eNt, (1)
где С - постоянная интегрирования.
Как видим, выражение (1) есть та самая экспоненциальная закономерность увеличения численности особей популяции, что соответствует мальтузианству в более поздней интерпретации. (В скобках замечу, что об экспоненциальном росте численности живых существ и, в частности, людей говорили и до Мальтуса). Из выражения (1), например, следует, что если в каком-нибудь заповеднике уничтожить всех волков, то оставшиеся без естественных врагов олени должны размножаться по экспоненциальной закономерности. В действительности же, как показывают наблюдения, рождаемость оленей, приходящуюся на одну самку, падает.
Теперь, если y = y(t) есть численность особей популяции хищников в момент времени t, а V(x) - закономерность, по которой единичный хищник уничтожает своих жертв, то скорость изменения численности особей популяции жертв будет:
![]() (2)
(2)
Таким образом, мы получили классический
вид первого уравнения математической модели Лотки-Вольтерры (см., например,
[5]).
Одним из распространненых видов уравнения (2) является:
![]() (3)
(3)
где M = const. Здесь подразумевается, что чем больше численность особей популяции жертв, тем больше они уничтожаются хищниками. Решая это уравнение, получим:
x = C e(N-M)t , (4)
где С - постоянная интегрирования.
Как видим, в выражение (4) входят три свободных параметра - N, M и C. Поэтому, если на каком-нибудь интервале времени t заданы значения x = x(t), то часто можно, скажем, методом наименьших квадратов подобрать такие значения параметров N, M и C, что выражение (4) будет довольно точно описывать закономерность изменения численности особей рассматриваемой популяции. Однако, это вовсе не означает, что полученным выражением можно будет также экстраполировать значения численности особей популяции за пределы заданного интервала времени t, т. е. математическая модель Лотки-Вольтерры не обладает прогностическим свойством, из-за чего биологи обычно говорят, что эта модель "хорошо работает лишь на бумаге".
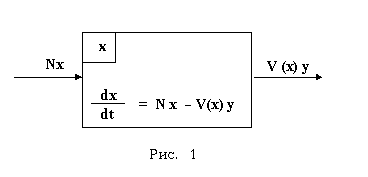
Теперь популяцию жертв из системы "хищник-жертва" рассмотрим как некоторую изолированную кибернетическую систему ("черный ящик"). Входом этой системы будет рождение новых особей, а выходом - гибель особей от деятельности хищников. И, если N = const есть средняя скорость размножения единичной особи популяции, то Nx будет скоростью размножения всей популяции. А если V(x) есть закономерность, по которой единичная особь хищников уничтожает своих жертв, то скоростью убыли численности особей популяции жертв будет V(x)y, где y есть число особей популяции хищников. Тогда уравнение (1) можно получить и без привлечения мальтузианского постулата (см. рис.1).
Следовательно, математическая модель Лотки-Вольтерры соотвествует такой гипотетической системе "хищник-жертва", в которой популяция жертв с точки зрения численности особей является изолированной кибернетической системой. А поскольку такого в природе не бывает, то в корне неверно не только мальтузианство, но вместе с ним и все те учения домальтусского и постмальтусского периодов, в которых постулируется способность живых существ размножаться экспоненциально при отсутствии гибельных факторов. И, конечно, вместе с ними неверна и математическая модель Лотки-Вольтерры.
Теперь, чтобы исправить указанную ошибку, рассмотрим популяцию жертв как некоторую кибернетическую систему с входом (рождение новых особей), выходом (убыль, гибель особей) и действующими на нее посторонними, зачастую случайными, факторами (наличие или отсутствие пищи, активность хищников, погодно-климатические условия, стихийные бедствия, болезни и т.д.). Также учтем, что популяции живых существ, как правило, не обмениваются своими особями с окружающей средой (гибель особей уже учтена), т.е. по отношению к численности особей всякая популяция живых существ является замкнутой кибернетической системой. Тогда, как утверждает принцип обратной связи кибернетики, такая система может продолжительно и устойчиво функционировать (а популяции живых существ, как известно, успешно живут и развиваются на продолжении сотен и тысяч лет) лишь при наличии отрицательной обратной связи между ее входом и выходом. Это, в свою очередь, означает, что при прочих равных условиях гибель одних особей популяции должна приводить к рождению других. На общебиологическом же языке получается, что на угрозу исчезновения, выражающуюся в насильственной гибели особей, всякая популяция живых существ должна отвечать вспышкой размножения. Следовательно, гибнущие живые существа должны испускать или излучать какие-то сигналы, которые стимулируют воспроизводительную функцию их остающихся в живых сородичей. (Несколько забегая вперед, отмечу, что, как установлено экспериментальными исследованиями, указанные сигналы гибнущие существа излучают, а не испускают, т.е., по всей вероятности, носителями указанных сигналов являются какие-то физические волны). И в результате всех этих рассуждений вместо рис.1 имеем рис.2, а вместо уравнения (2) - уравнение:
![]() (5)
(5)
где N, т.е. скорость размножения
особей популяции уже не постоянна, а зависит от активности хищников.
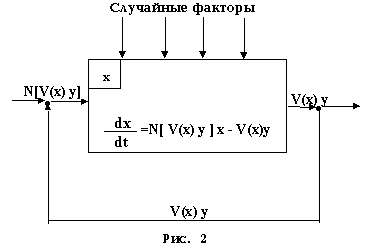
Остается только добавить, что уравнение (5) будет больше соответствовать действительности, если в ней под y = y(t) подразумевать не численность особей популяции хищников, а число реализаций всех тех факторов, которые в момент времени t приводят к гибели особей популяции жертв. В этом случае уравнение (5) будет действительно уже не только для систем "хищник-жертва", но и для всяких популяций всяких живых существ. И поскольку в основе уравнения (5) нет никакого постулата, то оно является уже не математической моделью, а точным математическим выражением, что расширяет возможности его применения.
Как было сказано, модель Лотки-Вольтерры была создана еще в 20-х годах нашего столетия. И, конечно, она много раз проверялась разными экспериментаторами. Тогда, почему они не заметили ту существенную ошибку, о которой говорилось? Или, почему не заметили, что гибнущие существа излучают сигналы, которые стимулируют воспроизводительную функцию их остающихся в живых сородичей? По очень простой причине: буквально во всех экспериментах по проверке уравнения (2) отсутствовал акт гибели. Например, опыты с лабораторными мышами проводились так: бралась размножающаяся группа животных и через определенные промежутки времени живым удалялось по одному выбранному жеребьевкой животному. Остальные продолжали размножаться. И все получилось как в уравнении (2). Я же, когда надо было проверять уравнение (5), предложил выбранного жеребьевкой животного сначала насильственно уничтожить в присутствии остальных, а затем удалить его труп. Контролем же предложил брать описанную схему по проверке уравнения (2), т.е. модель Лотки-Вольтерры.
Описанным путем уравнение (5) впервые проверялось в 1976-77 гг. на нелинейных белых лабораторных мышах в Новосибирском научно-исследовательском санитарном институте Министерства здравоохранения РСФСР. Опыты проводил известный экспериментатор, доктор биологических наук, профессор Сергей Сперанский. В результате с большой статистической достоверностью было доказано наличие искомого эффекта, что было опубликовано в 1978 г. [6] (подробности см. в разделе "Экспериментальные доказательства").
Таким образом, было сделано крупное научное открытие, которое в последующем вошло в некоторые учебники термодинамики политехнических институтов под названием "Эффекта Агаджаняна" (см., например,[7]).
Известно, что у многих биологических видов имеется половой диморфизм, который распространяется на морфологию, физиологию, этологию и другие параметры самцов и самок. И часто этот диморфизм до того велик, что самцы бывают с крыльями, а самки - без них (как, скажем, у некоторых видов насекомых), или самцы постоянно живут в воде, а самки - на суше (как, например, у одного вида африканских лягушек), т.е. самцы и самки одного и того же вида по разному будут реагировать на одни и те же внешние воздействия. Следовательно, всегда могут создаваться такие условия, когда только самцы или только самки одного и того же вида окажутся под угрозой. Т.е. биологические виды, представленные двумя полами, не смогли бы сколь-нибудь продолжительно жить и развиваться в нашем изменчивом мире. Однако, живут и развиваются. Тогда уже упомянутый принцип обратной связи кибернетики подсказывает, что эффект Агаджаняна также должен обладать половым диморфизмом. Т.е. половым диморфизмом должны обладать и предсказанные уравнением (5) сигналы гибнущих особей. А это означает, что гибель самцов должна приводить к преимущественному рождению самцов, а гибель самок - к преимущественному рождению самок. И это наблюдается у тех биологических видов (например, у некоторых видов пауков), самки которых уничтожают своих самцов сразу же после спаривания. Это подтверждается и тем, что во время крупных войн, когда гибнет много солдат-мужчин, мальчиков рождается больше.
С учетом сказанного рис.2
должен выглядеть как рис.3. Соответственно должно быть скорректировано
и уравнение (5).
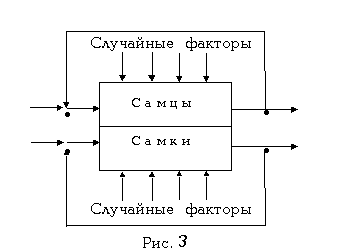
Хочу также отметить, что, по моему мнению, гибель является экстремальным случаем. Т.е. рассматриваемые здесь сигналы, пусть слабо, излучаются также при всяком стрессе и, вообще, всяком обмене веществ всеми органами и функциональными центрами, всеми клетками организма. Отсюда становится ясным, почему перед крупными войнами мальчиков рождается больше: тогда призываются в армию крупные мужские массы, которые в ходе подготовки к войне массово стрессируются. А их сигналы, суммируясь, приводят к рождению мальчиков. Или, другой пример: раньше в России говорили, что для того, чтобы родился мальчик, будущая мать должна слушать военные марши. Здесь дело, конечно, не в музыке, а в том, что раньше военные марши играли, как правило, при въезде войск в город и выезде из него. В это время опять приходили в движение крупные мужские массы и излучали "мужские сигналы". А наблюдавшие за этим процессом гражданские лица, в их числе и будущие матери, воспринимали эти сигналы. Дальше уже ясно.
Нечто аналогичное в недавнем прошлом наблюдалось и в Армении: во время разрушительного землятрясения 1988 года погибло много людей, а через девять месяцев был зафиксирован небывалый всплеск рождаемости, что, несомненно, явилось результатом гибели большого количества людей обоего пола. Затем к нам приехало много строительных бригад из всех республик СССР и, трудясь-стрессируясь день и ночь, эти крупные мужские массы стали восстанавливать разрушенное. В результате у нас, особенно в зоне бедствия, стали рождаться в основном мальчики. Некоторые демографы пытались это объяснить тем, что природа, якобы, восстанавливает потери нации. Однако, если бы это было так, то родилось бы больше девочек.
Долгие годы мне не удавалось опубликовть уравнение (5) - невозможно было получить соответствующее "добро". Все отмахивались от меня, бормоча: "Этого не может быть". И вдруг в 1985 году повезло: в подмосковном городе Пущино, в Центре билогических исследований Академии наук СССР был опубликован [8] вариант уравнения (5).Я даже получил приглашение выступить на Всесоюзном семинаре "Математические и вычислительные методы в биологии". Однако мое короткое выступление вызвало переполох у слушателей семинара, среди которых было немало тех, кто годами занимался математической моделью Лотки-Вольтерры.
Таким образом, обнаружено ранее неизвестное явление обратной связи между гибнущими и остающимися в живых особями одного и того же биологического вида на уровнях размножения и пола. Или, иначе, уравнение (5) действительно как для общей численности особей всякой популяции живых существ, так и для чисел их самцов и самок по отдельности. Однако, что такое ч и с л о ? Вещество? Энергия? Нет, информация.
Особи всякой популяции живых существ постоянно обмениваются веществом и энергией с окружающей средой, получая, скажем, вместе с пищей и возвращая вместе с выделениями, а после гибели или смерти также в виде составных частей их тела. Значит, по отношению к веществу и энергии (точнее, к указанному роду вещества и энергии) популяция является открытой кибернетической системой. Следовательно, те рассуждения, которые привели к получению уравнения (5), недействительны по отношению к этим двум компонентам. То же самое относится и к тому роду информации (например, заключенной в указанных веществе и энергии), которой популяция постоянно обменивается с окружающей средой. Однако есть и такая информация, которой особи популяции не обмениваются с окружающей средой. К этому роду относится информация о численности особей популяции, их половой принадлежности, жизненном опыте, признаках приспособления к гибельному фактору, иммунологическом статусе и т.д. И по отношению ко всей этой информации всякая популяция живых существ является замкнутой кибернетической системой. Следовательно, по отношению к ней можно применять все те рассуждения, которые привели к получению уравнения (5). Тогда вместо этого уравнения получим:
где H = H(t) - вся указанного рода информация популяции в момент времени t. Причем,
H = H1 + H2 + H3 + H4 + H5 + H6,
где H1 - информация,
определяющая факт гибели особей популяции;
H2 - информация, определяющая
пол гибнущих особей популяции;
H3 - информация, определяющая
жизненный опыт гибнущих особей популяции;
H4 - информация, определяющая
признаки приспособления к данному гибельному фактору;
H5 - информация, определяющая
иммунологический статус гибнущих особей популяции;
H6 - другая возможная информация,
по отношению к которой тоже популяция является замкнутой кибернетической системой;
y = y(t) - ччисло реализаций гибельных
факторов в момент времени t;
V =V(H) - закономерность, по которой гибнут
особи популяции.
Таким образом, между гибнущими и остающимися в живых особями популяции одного и того же биологического вида существует обратная связь на уровнях размножения, пола, жизненного опыта, признаков приспособления к данному гибельному фактору, иммунологического статуса и т.д. В последующем изложении по аналогии с уже принятым обозначением я каждый раз буду указывать о каком уровне эффекта Агаджаняна идет речь.